Video 01 de 11 - Sistema Imune -Teoria 01 - Biologia para Medicina - Landim
Fonte: http://www.youtube.com/watch?v=01QmEVLkDDE
CÉLULAS PROCARIÓTICAS
Os
organismos formados por células procarióticas designam-se por
procariontes, agrupam-se no Reino Monera e são conhecidos pela
designação geral de bactérias (do gr. : bakterion,
pequeno bastonete). Foram estes organismos que protagonizaram as
primeiras etapas da evolução dos seres vivos. Os registos
fósseis mais recuados datam de 3,46 milhões de anos. Nessa
época, a vida microbiana já era representada por
cianobactérias filamentosas, e a diversidade deduzida a partir
dos registos fósseis, conduziu a identificação de 11 taxons
diferentes.
No plano
morfológico, as bactérias apresentam uma enorme variedade de
formas e dimensões. Podem ainda viver isoladas ou formar
colónias de células mantidas agregadas depois de se dividirem.
Contudo, a despeito dessa variedade, é possível encontrar uma
unidade anatómica.
No que se
refere aos regimes de vida e ao metabolismo energético,
encontram-se bactérias que mantêm o regime heterotrófico
exclusivo, tal como certamente foram os primeiros seres vivos.
Outras, fotossintetizantes, são autotróficas, como as
bactérias verdes e as púrpura. Em ambos os grupos, existem
espécies sulfurosas (que oxidam o sulfureto de hidrogénio) e
outras que oxidam outras moléculas, mas nunca a água.
As
cianobactérias situam-se seguramente entre as mais recentes, na
escala evolutiva: o seu mecanismo de fotossíntese incorpora o
sistema que permite efectuar a oxidação da água, tendo como
consequência a libertação de oxigénio. Algumas das espécies
fotossintetizantes são também, cumulativamente,
heterotróficas. Algumas, mantêm-se obrigatoriamente
anaeróbias. Outras, pelo contrário, são aeróbias, e possuem
todo o equipamento enzimático que lhes permite efectuar a
respiração celular.
Anatomia da célula procariótica
As bactérias apresentam
uma extraordinária variedade de formas e a anatomia
ultrastrutural da célula reflecte a diversidade de regimes de
vida e de metabolismos energéticos. Seria fastidioso, no
contexto em que se inserem estas lições, enveredar por uma
descrição pormenorizada. Assim, abordaremos a anatomia da
célula bacteriana, descrevendo as principais estruturas
existentes nas diversas classes, referindo a função que lhes
assiste.
Quanto à forma que o corpo
celular pode apresentar, distinguem-se, grosso modo, quatro
modelos:
- Os cocos (coccus, cocci) são relativamente esféricos e formam, frequentemente, agrupamentos (esfilococos, em forma de cacho de uvas; estreptococos, em cadeia linear; sarcinas, em grupos compactos de oito; os diplococos (grupos de dois);
- Os bacilos (bacillus, bacilli) são ligeiramente alongados, com extremidades hemisféricas, podendo dispor ou não de flagelos;
- Os vibriões (vibrio) são encurvados, em forma de arco ou de vírgula, com um flagelo numa das extremidades;
- As espiroquetas são alongadas e helicoidais, podendo dispor de vários flagelos.são alongadas e helicoidais, podendo dispor de vários flagelos.
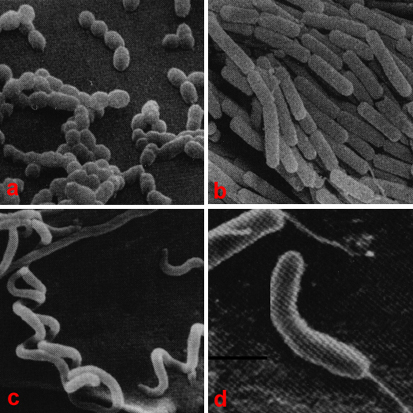 a: cocos; b: bacilos; c: espiroquetas; d: vibriões
a: cocos; b: bacilos; c: espiroquetas; d: vibriões
A célula bacteriana
possui, como qualquer célula viva, um genoma, um citoplasma e uma
membrana
plasmática. Com excepção dos
micoplasmas e das formas L., todas as bactérias possuem também
uma parede
celular. Algumas possuem ainda
uma cápsula externa.
Parede
celular
A parede celular, pela sua
rigidez, forma um estojo que estabiliza a forma característica
da célula, protegendo-a de agressões externas, nomeadamente das
variações de pressão osmótica.Com excepção das bactérias
halófilas, metanogénicas ou temoacidífilas (que se classificam
como Arqueobactérias), o composto principal da parede bacteriana
é o peptidoglicano. O peptidoglicano constitui assim o monómero de
uma densa rede macromolecular. É composto, ele próprio,pela N-acetilglucosamina
e pelo ácido N-acetilmurâmico, associados
a aminoácidos em quantidades variáveis.
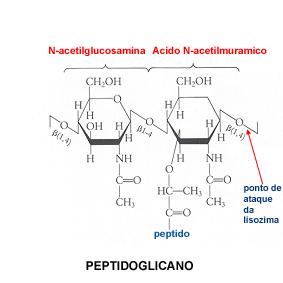
Esta estrutura de base da parede das bactérias
é mais ou menos importante e completada por constituintes
variáveis, mas específicos de cada espécie bacteriana. A
composição e a estrutura da parede celular determinam o
comportamento da célula face a um dos métodos de coloração
bacteriológicos: a
coloração de Gram.
Distinguem-se deste modo dois grupos principais de bactérias: as
bactérias gram-positivas, que se deixam corar pela coloração de Gram, e
as paredes das bactérias gram-negativas.
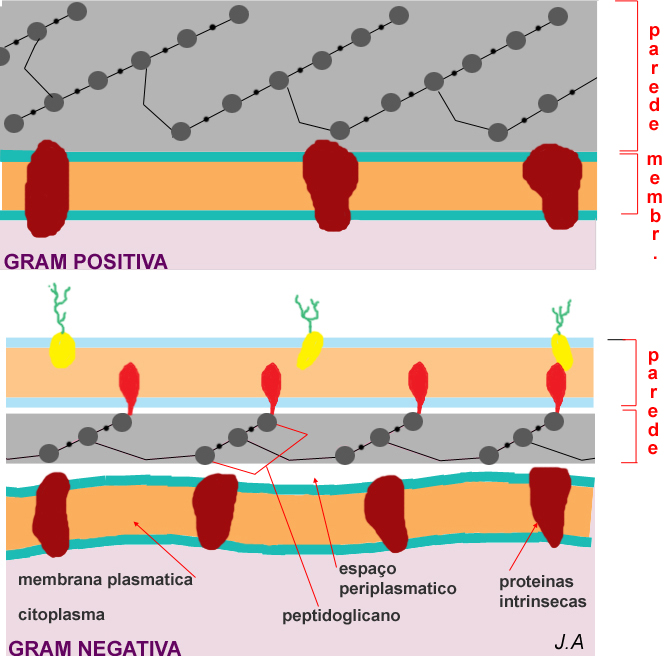
Esquema das paredes celulares em bactérias Gram positivas e Gram negativas
As bactérias gram-positivas possuem uma parede espessa e homogénea, ligada e encostada directamente à face externa da membrana plasmática. Nestes casos, não existe espaço periplasmático. A espessura destas paredes pode atingir 100 nm e representar até 30% do peso seco da célula.
Pelo contrário, a parede
das bactérias gram-negativas é formada por dois folhetos: o folheto
interno, constituído por uma delgada camada do
complexo de peptidoglicano (não excedendo 20 nm de espessura) e
não encostado à membrana plasmática; o folheto
externo, também designado por membrana
externa dada a sua estrutura ser semelhante à de uma
membrana unitária (constituída por liposacáridos e
lipoproteínas). A coesão entre os dois folhetos estabelece-se
através de lipoproteínas integradas no folheto externo e
ligadas por ligações covalentes a peptidoglicanos. No folheto
externo existem ainda canais proteicos que franqueiam a passagem
à água e a diversos metabolitos.
A diferença de
comportamento das duas paredes relativamente à coloração de
Gram reside essencialmente na técnica de coloração utilizada e
não na afinidade das duas paredes para o corante. Com efeito,
ambas as paredes são coradas pelo corante de Gram (violeta de
genciana e lugol). Contudo, no final, as células são lavadas
com um solvente (álcool ou acetona) que dissolve e elimina o
folheto externo das bactérias gram-negativas. Se bem que a
parede mucossacarídica subsistente seja suficientemente rígida
para garantir a integridade da célula, pela sua espessura
delgada, ela não retém suficientemente o corante. Pelo
contrário, as bactérias Gram positivas retêm o corante nas
suas espessas paredes.
A lisozima, enzima presente
em meios biológicos tais como secreções (lágrimas, saliva,
muco nasal, clara do ovo, etc.) ou no citoplasma das células
fagocitárias, hidrolisa o peptidoglicano e, consequentemente,
destroi a parede. Desprovidas de parede, as bactérias
transforma-se em protoplastos, extremamente vulneráveis às variações da
pressão osmótica.
A acção da penicilina
conduz ao mesmo efeito, não porque destrua o peptidoglicano, mas
porque inibe a sua síntese, durante o crescimento bacteriano.
A acção lítica da
lisozima ou a inibição de síntese do peptidoglicano pela
penicilina conduz, nas bactérias gram-negativas, à formação
de esferoplastos. Estes são equivalentes aos protoplastos das
bactérias gram-positivas, com a diferença que conservam o
folheto externo. Estas formas de bactérias tornadas deficientes
de parede podem, em cultura, reconstituir a parede. As formas
espontaneamente deficientes, designam-se por formas L.
Cápsula
Algumas
espécies bacterianas elaboram uma volumosa cápsula de natureza
polissacarídica (cuja espessura ultrapassa muitas vezes a
própria espessura da célula). Esta cápsula desempenha um papel
determinante na resistência à ingestão e à digestão pelas
células fagocitárias nos processos infecciosos, ou participa na
aderência dos organismos entre si ou ao substrato. É
particularmente o caso do pneumococus Streptococcus
pneumoniae.
Membrana
plasmática
A membrana plasmática
encontra-se encostada ou ligeiramente afastada da camada de
peptidoglicano pelo espaço
periplasmático. A estrutura
desta membrana é semelhante à das células eucarióticas,
registando-se contudo uma diferença na sua composição: o
colesterol, geralmente ausente nos procariontes, é substituído
por uma molécula semelhante a um esteroide, um hopanoide.
Na célula bacteriana, a
membrana é suporte de grande parte da actividade metabólica e
assegura, nomeadamente a cadeia da fosforilação oxidativa
(respiração, nas bactérias aeróbias). Para além disso,
assegura, como nas células eucarióticas o transporte selectivo
de moléculas e a saída dos enzimas responsáveis pela síntese
da parede celular.
Em algumas células, a
membrana forma pregas e invaginações que aumentam muito a sua
superfície, denominadas mesossomas.
Embora subsistam dúvidas sobre a função dessas estruturas,
formula-se a hipótese de serem zonas de intensa actividade
respiratória.
À superfície das
bactérias encontram-se filamentos proteicos longos, com 20 nm de
diâmetro. Uns são implicados nos processos de aderência das
bactérias entre elas ou a substratos; são designados por fímbria; outros, intervêm na transferência de material
genético entre bactérias; são designados por pili sexuais. Alguns autores designam ambos por pili (pilus,
pili)
Citoplasma
O citoplasma da célula
bacteriana ocupa todo o espaço intracelular. É um meio viscoso,
rico em proteínas, no qual se encontram numerosos ribossomas de
tipo 70S (semelhantes aos que se encontram nas mitocôndrias) e
inclusões de diversa natureza. Não contem nem organitos
membranares, nem vacúolos.
Entre as diversas
inclusões detectáveis no citoplasma, referem-se: os grãos de glicogénio e de poli-b-hidroxibutirato, que constituem reservas de carbono; os grãos de cianoficina, próprios das cianobactérias, que são
reservas de azoto sob a forma de aminoácidos; os carboxissomas, presentes em muitas cianobactérias e
bactérias nitrificantes e que são reservatórios de
ribulose-1,5-difosfato carboxilase, enzima específica do
mecanismo bioquímico de fixação do CO2;
os magnetossomas, partículas de magnetite e que proporcionam
orientação no campo magnético terrestre; ou ainda os vacúolos de gás, que são organitos de flutuação.
Genoma
O genoma bacteriano ocupa a
região central da célula, designada por nucleoide; é constituído por uma única molécula de
ácido desoxirribonucleico (ADN), bicatenária e fechada em anel,
o cromossoma bacteriano, comumente colado à membrana
plasmática. Em Escherichia coli, a
molécula de ADN tem 2 nm de diâmetro e 1,2 mm de comprimento.
Uma tão longa molécula (cerca de 500 vezes mais longa que a
própria célula), ocupa um espaço diminuto; para tal,
encontra-se profundamente enovelada.
O cromossoma não é o
único repositório de informação. Existem ainda pequenos
anéis de ADN, designados por plasmídeos,
geralmente todos idênticos, com autonomia de replicação
relativamente ao cromossoma. Os plasmídeos são responsáveis
por características específicas tais como (i)
a capacidade de fixação do azoto atmosférico, protagonizado
pelas bactérias do género Rhizobium,
(ii) a resistência a antibióticos, (iii)
a produção de toxinas ou ainda (iv) o
catabolismo de substâncias carbonadas como os hidrocarbonetos.
Os plasmídeos são transferíveis de uma bactérias a outras,
não só através da divisão celular, mas também através de
estruturas tubulares, os pili sexuais, que
estabelecem a ponte entre duas bactérias.
Membranas
fotossintéticas, clorossomas e tilacoides
A fotossíntese é uma
função existente em muitas bactérias, umas anoxigénicas
(que não libertam oxigénio) como as bactéria púrpura e as
bactérias verdes; outras oxigénicas (que produzem oxigénio), como as
cianobactérias. Todas possuem pigmentos fotorreceptores, mas as
estruturas em que estes se localizam são diferentes.
Nas bactérias púrpura, os pigmentos fotorreceptores (clorofilas
bacterianas a e b) encontram-se instalados em sistemas
membranares resultantes da hipertrofia e pregueamento da membrana
plasmática. Nas bactérias
verdes, os pigmentos (clorofilas
bacterianas a, c e outras) encontra-se localizados em
corpúsculos elipsoides designados por clorossomas. Embora se encontrem encostados à membrana
plasmática, os clorossomas são limitados por membrana
diferente da membrana unitária. Nas cianobactérias,
a clorofila (clorofila a) localiza-se em membranas sobrepostas
(dependentes da membrana plasmática ?) designadas por tilacoides, aos quais se associam partículas de
ficocianina, os cianossomas, onde se localizam os pigmentos do grupo das
ficobilinas.
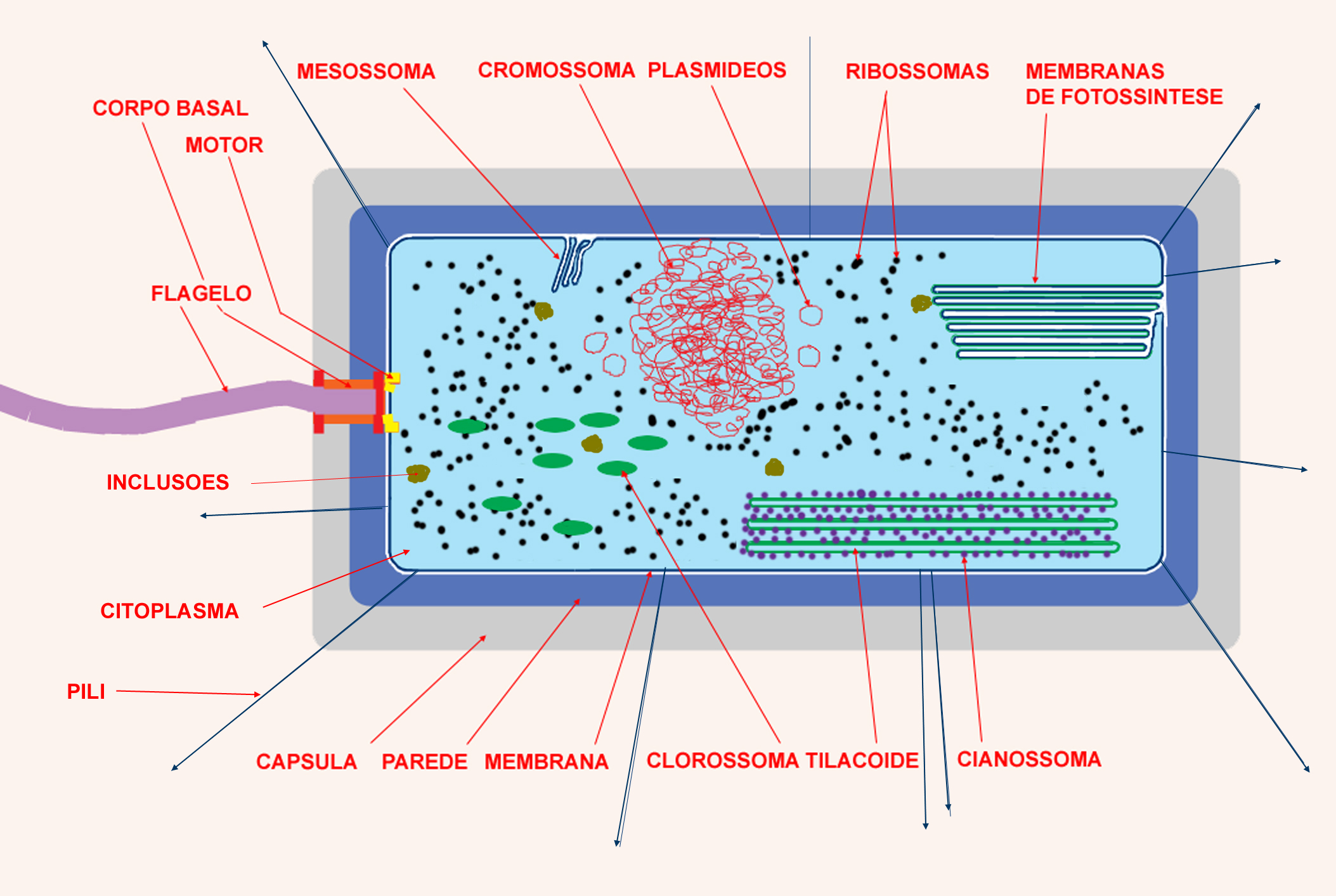
Esquema de uma bactéria hipotética que
reunisse em si todas as principais estruturas referidas
Flagelos
Sendo grande a diversidade
do Reino Monera, encontram-se bactérias que possuem
características muito diversas. Algumas são dotadas de
capacidade de locomoção. Para tanto, dispõem de um ou mais flagelos, com cerca de 20 nm de diâmetro. As
espiroquetas possuem um conjunto vasto de flagelos, enrolados
externa e helicoidalmente em volta da célula, denominados
flagelos periplasmáticos.
Os flagelos estão
ancorados através de uma estrutura que atravessa a parede
celular, o espaço periplasmático e a membrana plasmática,
designada por corpo basal. Sobre este corpo basal actua um motor
molecular, igualmente ancorado na membrana plasmática.
Fonte: http://materiais.dbio.uevora.pt/jaraujo/biocel/celulas.procarioticas.htm
Nenhum comentário :
Postar um comentário